近年、人為的要因によって自然環境の悪化が不可視的レベルでも急速に進んでいる。その要因の一つである化学物質は生体内においてホルモン様作用をもたらしたり、内分泌系に異常を引き起こすことが数多く報告されるようになってきた。特に、雄の雌化や性的異常を引き起こすホルモン作用をもつ化学物質や内分泌系に異常をもたらす化学物質、いわゆる内分泌かく乱化学物質の存在が解明されつつある。環境水を通して作用する化学物質の生体への影響を調査・確認するために、水生生物が指標動物として用いられ、その生殖異常が注目されている。この生殖系の異常を判定する評価基準としては、(1)形態的指標(生殖巣の組織変化、第二次性徴の消長)、(2)ビテロジェニン量、および(3)体重生殖巣重指数GSIが上げられている。
こうした環境要因の性徴への影響を調べるためには、遺伝的性と連関したマーカーをもつd-rR系(山本時男による育種系統)1)、S-rR系(住化テクノサービスによる育種系統)2)、あるいはFLF系(Qurt系統)3)のメダカを用いれば、雄に及ぼす女性ホルモン、雌に及ぼす男性ホルモンの作用をもつ環境要因の生殖系の分化・発達に及ぼす影響をより正確に評価・判定できる。
魚類には雌雄同体魚と雌雄異体魚がある。雌雄同体魚は、雌雄の生殖巣(腺)を分離型、あるいは混在型の形態で同時にもち、生涯の時期や生態的環境によって多様な発達の変化を示す4)。すなわち、生理的な雌性先熟や雄性先熟の雌雄同体魚類において、それぞれの生殖巣は生涯の時期、社会的、生態的や季節的な要因、あるいは化学物質など種々の要因によって性の変換(sex succession)5)を引き起こすため、非常に複雑である。したがって、内分泌かく乱化学物質の影響を調べるのに、雌雄同体魚を指標動物として扱うのは必ずしも適切ではない。一方、雌性生殖巣、あるいは雄性生殖巣のいずれかしかもたない雌雄異体においては、一たん性分化すると生涯その性が変換することのない“性の分化種”と生涯の時期によって変換する“性の未分化種”がある。前者の性の分化種であれば、生殖巣の変化をその分化時に外的要因によって影響を受けたものと捉えることが可能である。
精巣卵:
花岡(1947)6)によれば、両生類において実験的に精巣に卵を最初に誘導した人はMeynsで、1920年代以後精巣卵の誘導実験が数多く報告されている。性の分化型であるメダカにおいても、精巣のシスト内に二次精原細胞、精原細胞の正常分裂、あるいは異形分裂(heterotypic division)の前期に一つの異常発達した核小体をもつ精原細胞、もしくは精母細胞から小さい偽(擬似)卵母細胞(pseudo-oocytes)が生じることが早くから知られている7)。その他、性ホルモン(サケ科 8-10), グッピー11,12))とか、他のステロイド系物質あるいはX線照射(メダカ13,14))によって成魚の精巣内に卵母細胞が生じることも報告されている。メダカでは、自然状態においては雄魚を調べても性転換、精巣卵の形成は殆ど観察されない15,16)。しかし、実験的に雄成魚をエストロジェロンで処理すると、処理後2週間以内に成魚の精巣内には巨大精原細胞が出現し、3週間以内に精巣内に卵様細胞が出現することを小川(1959)17)もみている。精巣内の卵様細胞はシスト内に存在し、すべて発達が同調している。しかも、エストラジオールで処理し続けると、精巣内の卵母細胞は退化するし、精子形成も抑制される18,19)。
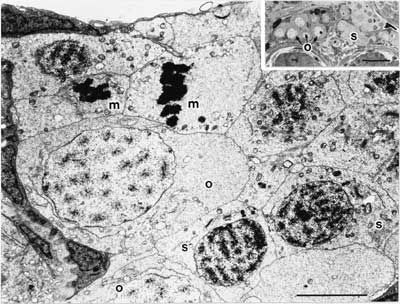
成魚をエストラジオールで処理すると、6日目の精巣にザイゴテン期の卵母細胞が認められ、パキテン期およびデプロテン期のものは12日目に現れる19,20)。柴田・濱口19)によれば、タイプA精原細胞は、それぞれが接着したセルトリ細胞によって仕切られている。タイプA精原細胞が分裂して、セルトリ細胞に接着しないままで増殖するタイプB精原細胞が生じる。セルトリ細胞によって形づくられたシスト内で、タイプB精原細胞は減数分裂に移る前に9〜10回分裂する。1つの精巣内のシスト中に形成された卵母細胞はその数から、4〜5回分裂した段階のタイプB精原細胞から生じるものと判断されている。電子顕微鏡写真(図1)は、精細管中のシストの中に集団で存在している形成初期の卵母細胞である。
 |
|
図1 エストラジオール-17βの投与によって生じた精巣卵(体長9 mm)
|
これまで、メダカの精巣中に生じた卵母細胞は、卵母細胞様細胞(oocyte-like cells7))、卵様細胞17)(“egg-like”cells18))、卵形細胞 (“oviform” cells1)、oval cell21))、精巣卵(testis-ova13,14,20))などと記載されており、その和名および英名は統一されておらず、研究者によって異なる。近年、環境水における内分泌かく乱化学物質による生殖巣の異常、とくに雄性生殖巣内に卵母細胞の形成(精巣卵の出現)に関する報告が数多くなされている。これらの報告書には、精巣中に生じた卵母細胞に対して精巣卵(testis-ova)という用語を当てて記述しており、そうした扱いが研究者間に定着しているようである。
卵精巣:
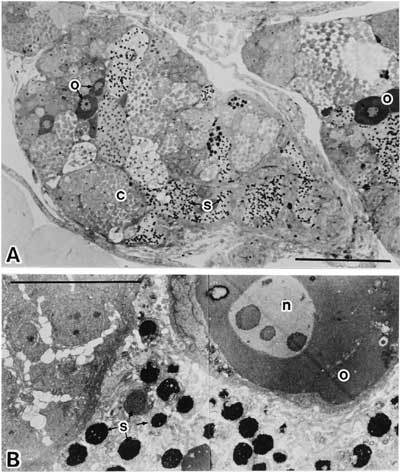
他方、山本1)は遺伝的雌を男性ホルモンで処理すると、雄性化することを証明した。“性の分化種”であるメダカの生殖巣原基には両性能(両性的生殖性:ambisexuality, bisexual potency)があり、遺伝的に反対の性にも転換することが可能であることを示した。グッピーでも、雌の未分化生殖巣は男性ホルモン処理によって遺伝的雄の生殖巣と同じ組織学的形成過程を経て精巣へと分化・発達する22)。男性ホルモン処理を受けたメダカにおいて、完全な雄への転換が起きず、卵母細胞の間に精子形成を部分的に起こしている遺伝的雌の生殖巣がしばしば見られる。卵巣に部分的に精巣要素をもつモザイク生殖巣である(図2)。そのモザイク生殖巣をどのように表現するのが適切であろうか。ニワトリ雌では、一方の卵巣を除去すると、もう一方の生殖巣(卵巣)にovotestisが生じると述べている23)。また、ovotestisの表現はvan Tienhoven(1983)24)によっても進化の途中にovotestisの段階を経由して卵巣から精巣になるという例のところで用いられている。このとき以外のXX、あるいはモザイクXX/XYの間性の表現型にもovotestisを用いている。アンブリストーマの生殖巣移植実験においては、卵巣が卵を含む精巣組織になったものをovotestisと記載している25)。最近、コイXXの劣性突然変異masミ1(雄性化遺伝子)をもつ個体の中に卵母細胞と精母細胞が混在する生殖巣をもつものがあり、この生殖巣にもovotestisという用語が当てられている26)。生物学辞典27)を見れば、「哺乳類などの間性(intersex)で卵巣中に若干の精巣組織を混じている場合、これを卵精巣ovo-testisということがある」と記述している。これらは総じて、遺伝的雌の生殖巣(予定卵巣)が一部に精母細胞や精子をもつ精巣組織への変化した生殖巣に対して卵精巣ovotestisという表現をしている。しかし、エゾサンショウウオにテストステロン・プロピオネートを注射した場合、「精巣から変型して生じたと思われる組織の内にも異常に大きい卵細胞が発達していわゆる卵精巣(ovotestis)を作ることもある」25)と述べている場合がある。
 |
|
図2 メチルテストステロンの投与によって生じた卵巣精子(体長23 mm)
|
また、医学辞典をみると、卵精巣の記載はあるが、精巣卵については全く書かれていない。遺伝学辞典28)においても、精巣卵の記載はないが、卵精巣(ovotestis)について「(1)雌雄同体の動物である軟体動物腹足類(カタツムリなど)でみられるように、先に精子をつくり、続いて卵をつくったり、あるいは同一器官内の一部で精子を、一部で卵をつくったりするような生殖器官をいう。 (2)動物の卵巣の中に一部精巣組織が混じったもので、間性でみられる。」と解説している。この場合、卵精巣は必ずしも遺伝的雌において卵巣の一部に精子、あるいは精巣組織が生じるというとらえ方をしていない。ドーランド図説医学大辞典29)にも、ovo-testisを「卵巣睾丸」と書いてあり、「精巣と卵巣の両組織をもつ生殖腺」と遺伝的性に関係なく、両性的表現がなされている。ovo-testisとは「ovum-testis」のイメージを受ける。そのため、「精巣内の卵」でtestis-ovumと区別するのが難しくなる。卵巣内に精巣組織が生じたものとは限らない表現である。
以上のように、遺伝的雄の生殖巣(精巣)内に存在する卵を指して精巣卵と呼んでいるのに対して、卵精巣(ovotestis)は必ずしも遺伝的雌の卵巣内に存在する精子、あるいは精巣組織を指しての表現とは限らないものがある。精巣卵に対応させる意味で、遺伝的雌の生殖巣(卵巣)内に卵母細胞と混在する精子(あるいは精巣組織)に対しては、既存の用語である卵精巣(ovotestis)よりも、むしろ卵巣精子(ovary-sperm)と表現することを提案したい。
引用文献
1) Yamamoto, T. (1958) J. Exp. Zool., 137: 227-264.
2) Hagino, S., M. Kagoshima and S. Ashida (2001) Environm. Sci., 8: 75-87.
3) Wada, H., A. Shimada, S. Fukamachi, K. Naruse and A. Shima (1998) Zool.
Sci., 15: 123-126.
4) 中園明信 (1991) 魚類生理学(板沢靖男・羽生功 編),恒星閣厚生社.
5) 鈴木克美 (1989) 栽培技研,18: 45-55.
6) 花岡謹一郎 (1947) 生物,増刊1号,146-161.
7) Oka, T.B. (1931) J. Fac. Sci. Imp. Univ. Tokyo, IV, 2: 219-223.
8) Padoa, E. (1937) Monit. Zool. Ital., 48: 195.
9) Padoa, E. (1939) Bio-Morphosis, 1: 337.
10) Baldwin, F.M. and M.H. Li (1945) Amer. Nat., 79: 281.
11) Berkowitz, (1938) Anat. Rec., 71: 161-175.
12) Berkowitz, (1941) J. Exp. Zool., 87: 233-243.
13) Okada, Y.K.(1943) Proc. Imp. Acad. Tokyo, 19: 501-504.
14) Okada, Y.K.(1944) Proc. Imp. Acad. Tokyo, 19: 244-250.
15) Aida, T. (1921) Genetics, 6: 554-573.
16) Yamamoto, T. (1953) J. Exp. Zool., 123: 571-594.
17) 小川嘉一郎(1959) 動物学雑誌,68:159-168.
18) Hishida, T. and H. Kobayashi (1985) J. Liber. Art. Asahi Univ., 11: 95-111.
19) Shibata, N. and S. Hamaguchi (1988) J. Exp. Zool., 245: 71-77.
20) Egami, N. (1955) Jap. J. Zool., 11: 21-33.
21) 畠山 仁・糸沢佳津子・川俣明日香・原田隆彦 (1999) 日本獣医畜産大学研究報告,48: 50-56.
22) 宮森弘子 (1964) 実験形態学雑誌,18: 30-39.
23) Nalbandov, A.V. (1964) Reproductive Physiology. 2 nd Ed., W.H. Freeman
and Company, pp. 316, San Francisco and London.
24) van Tienhoven, A. (1983) Reprodective Physiology of Verebrates. 2nd Ed.,
Cornell Univ. Press, pp.491, Ithaca and London.
25) 花岡謹一郎 (1954) 性(岡田 要・木原均編),pp.167-218,共立出版.
26) Komen, J., M. Yamashita and Y. Nagahama (1992) Develop. Growth Differ.,
34: 535-544.
27) 山田常雄・前川文夫・江上不二夫・八杉竜一 (1966) 生物学辞典,第3版,岩波書店(東京).
28) 田中信徳 (1977) 遺伝学辞典,pp.576,共立出版.
29) ドーランド医学大辞典編集委員会 (1998) 第28版 ドーランド図説医学大辞典,廣川書店.