■ニューロステロイドの発見
生体内のステロイド合成細胞では、コレステロールをもとにチトクロームP450scc(P450分子種の一つ;コレステロール側鎖切断酵素)の作用によりプレグネノロンがつくられる。このプレグネノロン合成が生体内に存在する全てのステロイドの生合成の第一段階であり、脳におけるステロイド生合成を証明するには、プレグネノロンの合成をまず明らかにする必要があった。通常は血液−脳関門を通過できない親水性の高いプレグネノロン硫酸エステルがラットの脳に高濃度に存在することに着目したBaulieuらの研究により、脳におけるプレグネノロンとその硫酸エステルの合成がラットで見いだされた[総説1]。一方、我々はウズラやカエルなどの脳細胞ミトコンドリアとコレステロールを反応させて生化学的に解析した結果、チトクロームP450sccがこれらの脳細胞に存在してプレグネノロンが合成されることを明らかにした[総説2-3]。
■ニューロステロイドの生合成
その後の研究により、チトクロームP450sccに加えてステロイド硫酸基転移酵素(HST)、3β-水酸基脱水素/Δ5-Δ4-異性化酵素(3β-HSD)、5α(β)-還元酵素、チトクロームP45017α,リアーゼ(P450分子種の一つ;17α-水酸化酵素・開裂酵素)、17β-水酸基脱水素酵素 (17β-HSD)など多くのステロイド合成酵素が脳に存在することが証明され、脳はコレステロールをもとにプレグネノロンとプレグネノロン硫酸エステルを初めとするさまざまなニューロステロイドを合成していることが明らかになった[総説1-3]。ニューロステロイドは末梢内分泌腺を除去してもあまり変動しないことから、末梢内分泌腺とは独立したステロイド合成系が脳に存在する。
脳におけるニューロステロイド生合成経路の大略を我々は鳥類の研究から次のように明らかにした[総説4]。まずチトクロームP450sccによりコレステロールからプレグネノロンが合成される。ステロイド硫酸基転移酵素(HST)の作用により一部のプレグネノロンはプレグネノロン硫酸エステルとなる。さらに3β-HSDによりプレグネノロンから性ステロイドとして知られるプロゲステロンが合成される。プロゲステロンは5β-還元酵素と3β-水酸基還元酵素(3β-HSO)により3β,5β-テトラハイドロプロゲステロン(エピプレグナノロン)に代謝される。哺乳類の脳では5α-還元酵素と3α-水酸基還元酵素
(3α-HSO) によりプロゲステロンは3α,5α-テトラハイドロプロゲステロン(アロプレグナノロン)に代謝される。両生類や魚類の脳でもプレグネノロンやプロゲステロンなどのニューロステロイドが合成される。一方、脳にはチトクロームP45017α,リアーゼが存在しており、プロゲステロンから17α-ヒドロキシプロゲステロンを経てアンドロステンジオンが合成される。鳥類の脳には17β-HSDも存在しており、アンドロステンジオンからアンドロゲンであるテストステロンが合成される。脳におけるチトクロームP450arom(P450分子種の一つ;芳香化酵素)の存在は古くから知られており、脳はさらにアンドロゲンからエストラジオール、エストロンなどのエストロゲンを独自に合成する。
■ニューロンのニューロステロイド生合成
ニューロステロイドの作用を解析するには、ニューロステロイドを合成する細胞を同定することが不可欠である。Baulieuらの研究から、まずオリゴデンドロサイトやアストロサイトなどのグリア細胞によるニューロステロイドの合成が証明された[総説1]。我々は小脳の皮質ニューロンであるプルキニエ細胞がニューロステロイドを合成することを見いだし、ニューロンによるニューロステロイド合成を明らかにした[総説3]。
我々のウズラを用いた免疫組織化学的解析と生化学的解析により、小脳皮質ニューロンであるプルキニエ細胞にチトクロームP450sccが密に局在していることが見いだされた[5]。ラット[6]やカエル[7]でも同様であり、プルキニエ細胞が脳の代表的なニューロステロイド合成細胞であることは脊椎動物に一般化される重要な発見である。さらに、我々はラットを用いたin
situ hybridization法による解析により、プルキニエ細胞にはチトクロームP450sccに加えて3β-HSDが局在していることを見いだした[8]。興味深いことに、このニューロンではチトクロームP450sccの発現は生後から成熟期にかけて恒常的に認められるが、3β-HSDの発現は新生期に増加する[6,
8]。さらに生化学的に解析したところ、プルキニエ細胞では生後から成熟期にかけて恒常的にプレグネノロンとその硫酸エステルが合成されるが、新生期にはプレグネノロンからプロゲステロンとその代謝ステロイドであるアロプレグナノロン
(3α,5α-テトラハイドロプロゲステロン) の合成が高まることが明らかになった[6, 8]。
■ニューロステロイドによる樹状突起伸長とシナプス形成の誘導作用
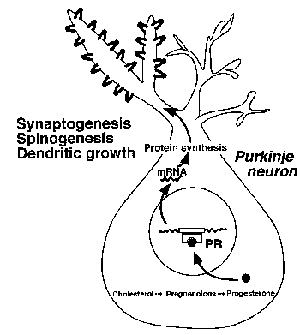
小脳の皮質は生後まもない新生期に形成される。この時期の小脳ではニューロンの発達やシナプス形成が活発になされ、小脳の神経回路が構築される。まず、新生期のプルキニエ細胞が活発に合成するプロゲステロンの作用を、ニューロンの発達とシナプス形成に着目し、超微形態学的に解析した。出生直後のラットを用いた解析から、プロゲステロンにはプルキニエ細胞の樹状突起の伸長を導く作用があることが明らかになった[9]。樹状突起の形態を詳しく解析したところ、プロゲステロンは樹状突起の棘形成と棘シナプス形成を誘導することも見いだされた[9]。我々は、プルキニエ細胞の樹状突起の伸長、樹状突起上の棘と棘シナプスの形成を誘導するプロゲステロンの作用機構を明らかにするために、プロゲステロン受容体の同定をおこなった。その結果、プルキニエ細胞の核内にはプロゲステロン受容体
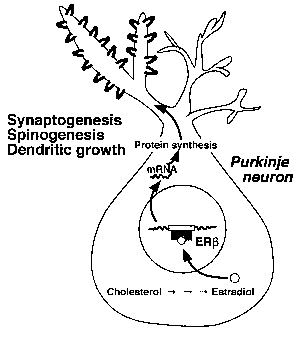
(PR-AとPR-B) が局在しており、プロゲステロンはこれらのPRを介したゲノミック作用により、樹状突起の伸長、樹状突起上の棘シナプスの形成を導くことが明らかになった[9](図1)。さらに、最近の我々の研究により、新生期のプルキニエ細胞ではチトクロームP450aromが発現しており、プロゲステロンに加えてエストラジオールが合成されることが見いだされた[10]。詳しい解析の結果、エストラジオールにもプルキニエ細胞の樹状突起の伸長を誘導する作用があることが明らかになった[10]。プルキニエ細胞の核内にはエストロゲン受容体(ERβ)が局在しており、エストラジオールはERβを介したゲノミック作用により樹状突起の伸長を誘導することがわかった(図2)。
以上のプロゲステロンとエストラジオールのゲノミック作用により、新生期には小脳の機能を担うハードウェアである神経回路が構築されると考えられる[総説11]。プロゲステロンとエストラジオールは卵巣が合成する性ステロイドとして知られているが、新生期の卵巣ではプロゲステロンとエストラジオールの合成は低く、プルキニエ細胞が独自に合成するプロゲステロンとエストラジオールがニューロンの発達とシナプス形成を誘導するという大変に重要な発見がなされた。
 |
 |
|
図1
|
図2
|
■ニューロステロイドによるシナプス情報伝達調節作用
従来、生体内のステロイドは標的細胞内(多くは核内)に存在する受容体を介したゲノミック作用により脳機能を調節すると考えられていたが、ある種のニューロステロイドは急性的なノンゲノミック作用によりシナプスの情報伝達を調節することがわかった。我々はプルキニエ細胞が合成するニューロステロイドであるプレグネノロンとプレグネノロン硫酸エステルの作用をラットの小脳スライスを用い電気生理学的に解析した。パッチクランプ法によりプルキニエ細胞からシナプス電流の発生頻度を調べたところ、プレグネノロン硫酸エステルには抑制性のシナプス電流の発生頻度を増加させる作用があることがわかった。この効果は急性的であり、プレグネノロン硫酸エステルの潅流を開始してわずか数分後から検出された。詳しい解析の結果、シナプス電流の発生頻度を急性的に増加させるプレグネノロン硫酸エステルの作用機序は次のように要約された。(i)プルキニエ細胞で合成されたプレグネノロンはステロイド硫酸基転移酵素(HST)によりプレグネノロン硫酸エステルとなる。(ii)プレグネノロン硫酸エステルはプルキニエ細胞から傍分泌されてプルキニエ細胞に投射するGABAニューロンに作用する。(iii) GABAニューロンの活動が高まり、シナプスにおけるGABAの放出頻度が増加する。(iv)その結果、プルキニエ細胞の活動が変化する。このように、プレグネノロン硫酸エステルにはシナプス情報伝達を調節する情報伝達調節因子としての作用があり、細胞膜にある受容体を介したノンゲノミック作用である、と結論された[総説3, 総説11]。
■まとめ
脳はコレステロールをもとにさまざまなニューロステロイドを合成する。小脳皮質ニューロンであるプルキニエ細胞は脳の代表的ニューロステロイド合成細胞であり、この細胞を実験系とした我々の研究により、脳におけるニューロステロイドの合成と作用の理解が得られた。ニューロステロイドにはニューロン樹状突起の伸長、シナプス形成、神経回路構築などを促進するゲノミック作用と構築された神経回路のシナプスにおける情報伝達を調節するノンゲノミック作用があることが見いだされた。
本研究は文部科学省科学研究費補助金(特定領域研究A、C;基盤研究A、B)と科学技術振興機構(CREST)の援助により得られたものであり、浮穴和義、坂本浩隆、松永昌宏、食見花子、臼井真理子、古川康雄、河内千恵、高瀬稔、山崎岳、小南思郎、R.W.
Lea諸氏との共同で行われた。
文 献
[1] Baulieu, E.E. (1997). Rec. Progr. Hormone Res. (review)
52, 1-32.
[2] Tsutsui, K. et al. (1999). Comp. Biochem. Physiol. C (review) 124, 121-129.
[3] Tsutsui, K. et al. (2000). Neurosci. Res. (review) 36, 261-273.
[4] Tsutsui, K. et al. (2003). Avian Poultry Biol. Rev. (review) 14, 63-78.
[5] Usui, M. et al. (1995). Brain Res. 678, 10-20.
[6] Ukena, K. et al. (1998). Endocrinology 139, 137-147.
[7] Takase, M. et al. (1999). Endocrinology 140, 1936-1944.
[8] Ukena, K. et al. (1999). Endocrinology 140, 805-813.
[9] Sakamoto, H. et al. (2001). J. Neurosci. 21, 6221-6232.
[10] Sakamoto, H. et al. (2003). Endocrinology 144, 4466-4477.
[11] Tsutsui, K. et al. (2003). J. Steroid Biochem. Mol. Biol. (review), 85,
311-321.